Cardiocondyla
Cardiocondyla is an Old World genus of myrmicine ants, comprising 48 described species, with an estimated total of ~100 species. The common name ‘heart-node ants’ refers to the shape of the post-petiole, which is usually broader than the petiole. Cardiocondyla are small ants (1.5 – 3 mm worker body length), have a three-segmented antennal club, propodeal spines, and a slender, elegant body shape.
Like related Nesomyrmex and Xenomyrmex, Cardiocondyla typically have small colonies of a few dozen to a few hundred workers. The majority of species nests in the soil, while several species of the “C. argentea” clade nest under rocks or in rock-crevices, and species of the C. wroughtonii clade have adopted an arboreal lifestyle, inhabiting ephemeral small nest sites under bark and in hollow twigs. The workers usually forage singly for small food items and fluids, and recruit nestmates by tandem running. Due to their potent venom, they are ignored by most larger ant species such as Pheidole and Solenopsis.
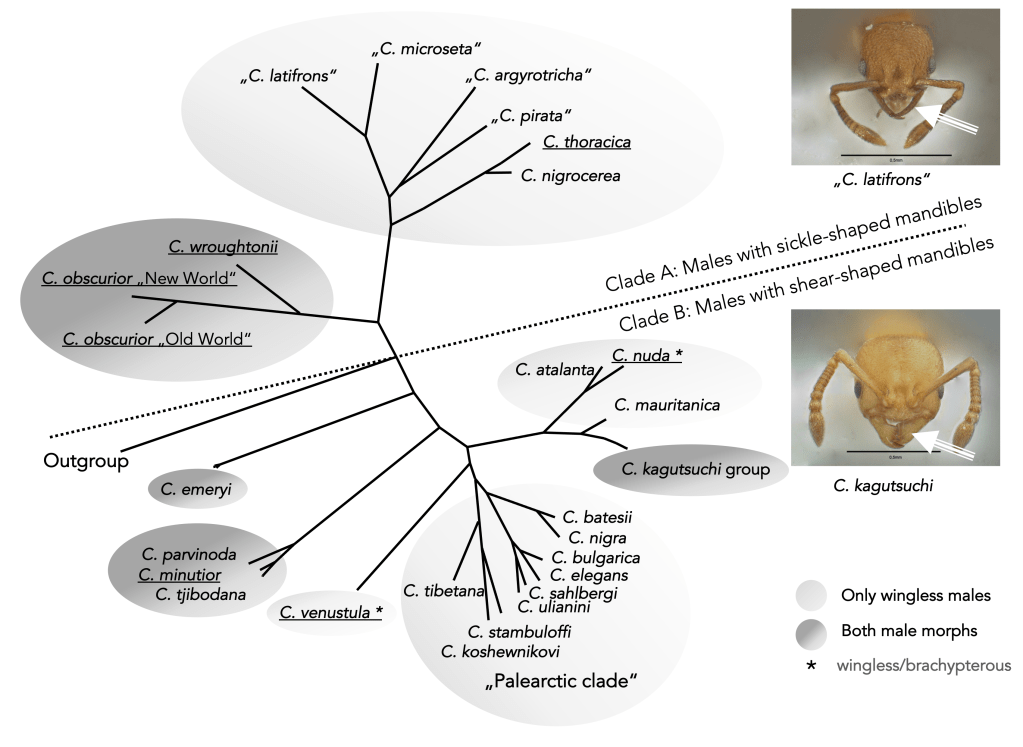
Several Cardiocondyla species are found in warm climates around the globe, and can be kept easily in the lab. These species represent both clades within the genus, which are characterized by the morphology of the mandibles of the wingless males (see phylogenetic tree below). Cardiocondyla is pre-adapted to a tramp lifestyle because of intranidal mating of wingless males with closely related females, thus allowing formation of new populations from tiny propagules.
Significance for evolutionary biology
Cardiocondyla ants are characterized by a wide range of evolutionary innovations, such as wingless males which fight until death to monopolize matings with nest mate queens. Wingless males facilitate mating between close kin and have accelerated the evolution of the genus. Cardiocondyla females display an equally fascinating diversity of traits, ranging from independent evolution of monogyny from the ancestral polygynous state in two lineages to oedipal (mother-son) mating in at least two species, genetic and maternal caste determination, and variation in mating frequency across the genus.
Cardiocondyla obscurior
The biology of one species in particular has been studied extensively. It has the smallest genome of any ant known so far (~190MB), and serves as model for e.g. the study of aging (Oettler and Schrempf 2016; Harrison et al. 2021; Jaimes-Nino et al. 2022, Jaimes-Nino & Oettler 2025, Jaimes-Nino et al. preprint), adaptation (Schrader et al. 2014, Errbii et al. 2021, Errbii et al. 2024, ), developmental plasticity (Schrader et al. 2015; Klein et al. 2016b; Schrader et al. 2017; 2021, Oettler et al. 2019, Schultner et al. 2023, van den Bos et al. 2024, Brülhart et al. 2024, Oettler et al. preprint), and immunity (Ugelvig et al. 2010; Schrempf et al. 2015; Wyschetzki et al. 2016, Leponiemi et al. 2021). The ant hosts an enigmatic endosymbiont, Candidatus Westeberhardia cardiocondylae (Klein et al. 2016a, Jackson et al. 2022), which also has a tiny genome (~533kb, 372 protein-coding genes). The ancestral Old World lineage of C. obscurior carries a manipulative Wolbachia strain, which is replaced in the New World by a second, docile Wolbachia strain (Ün et al. 2021, 2022). A 3D atlas of the worker brain is available (Bressan et al. 2014), and it is a good model to study worker task performance and maturation at the behavioral and molecular level (Oettler et al. 2015).